Převzato se svolením autora ze sborníku Botanický systém a evidence rostlin v botanických zahradách, ISBN 978-80-903697-3-3. Tento příspěvek je zkrácenou a pro tisk upravenou verzí přednášky přednesené na semináři Unie botanických zahrad dne 22. května 2007.
Úvod
Fylogenetická klasifikace cévnatých rostlin procházela v druhé polovině minulého století relativním obdobím klidu. U krytosemenných rostlin se ustálily klasifikace „koryfejů“ systematiky angiosperm, založené primárně na morfologických a dále anatomických, karyologických a palynologických znacích; u vyšších taxonomických kategorií (podtřídy, řády) byl kladen značný důraz na znaky fytochemické, zejména tvorbu různých sekundárních metabolitů. Anglosaská literatura používala nejčastěji klasifikaci Cronquistovu (Cronquist 1981, 1988), středoevropská literatura včetně našich učebnic (Hendrych 1977, 1986) inklinovala k pojetí Tachtadžjanovu (Tachtadžjan 1977); obě si však byly značně podobné. Pro jednoděložné rostliny vyšla pak koncem osmdesátých let zásadní příručka (Dahlgren, Clifford, Yeo 1985) v níž bylo užito Dahlgrenovo schema příbuznosti podle tzv. dahlgrenogramů; tato klasifikace byla posléze v mnoha bodech potvrzena i molekulární systematikou. Československá botanika byla ovlivněna i zásadní příručkou Novákovou (Novák 1961, 1972), jejíž informatická hodnota na úrovni charakteristik čeledí byla vysoká, avšak systém vyšších kategorií, založený na některých paleobotanických teoriích počátku 20. století, se neujal.
Kladistika a molekulární systematika
Převrat v nazírání na fylogenetický „strom“ nastal zprvu na úrovni metodologické, stále větším prosazováním kladistiky, jejímž úhelným kamenem je princip striktní monofylie: zjednodušeně řečeno, monofyletický taxon v sobě zahrnuje všechny potomky společného (hypotetického) předka. Tím se liší od evoluční (fyletické) klasifikace, na níž jsou založeny všechny výše citované práce; tato klasifikace uznává jak taxony monofyletické, tak parafyletické (parafyletický taxon vznikl ze společného předka, nezahrnuje však všechny jeho potomky, společného předka s ním mohou mít i další taxony). Teorii kladistiky publikoval v r. 1950 německý zoolog Willi Hennig, do širšího povědomí, zejména zoologické vědecké obce, však vstoupila až po překladu jeho práce do angličtiny (Hennig 1966).
V botanice se začala kladistika prosazovat koncem osmdesátých let 20. století a předcházející systémy pak zcela vytlačila s nástupem molekulární systematiky v botanice, za jejíž „starting point“ můžeme považovat rok 1993, kdy vyšla první publikace srovnávající molekulární příbuznost 499 druhů krytosemenných (Chase et al. 1993). Molekulární systematika využívá molekulárních dat z genových sekvencí určitých úseků DNA resp. RNA. Výhody molekulárních znaků tkví zejména v tom, že (1) molekulárních dat (=znaků) je řádově více než jakýchkoliv jiných, (2) jsou jednoduše a jednoznačně interpretovatelné (adenin je vždy adenin); ne zcela se potvrdilo jedno z původních tvrzení molekulární systematiky, že změny na úrovni genů podléhají méně konvergenci či paralelismu. Nejčastěji užívané genové sekvence přináší následující tabulka:
_____________________________________________________
Tab. 1 – Nejčastěji užívané genové sekvence
• Chloroplastová DNA
rbcL – kóduje podjednotku fotosyntetického enzymu RuBisCO; atpB, ndhF, trnK
• Nukleární ribosomální RNA
18S – 26S
• Mitochondriální DNA
atp1, atpA – pomalá evoluce, spíše pro původ krytosemenných
_____________________________________________________
Zpočátku byly vytvářeny fylogenetické stromy jen na základě (byť čím dál tím většího počtu) jednoho či více souborů molekulárních dat, koncem 20. století se pak objevuje řada prací, pokoušejících se o interpretaci na základě srovnání molekulárních a „morfologických“ dat (např. Nandi 1998); tento proud vyústil v souborné dílo pod redakcí manželů Soltisových (Soltis et al. 2005). Termín morfologický znak je zde užíván v širokém slova smyslu jako znak interpretovatelný (pozorovatelný) na jiné úrovni než molekulární (tj. včetně např. fytochemie, karyologie apod.) a jako protiklad „molekulárního“ znaku.
Význam molekulárních dat pro ilustraci vzájemných fylogenetických vztahů se projevil od počátku, byť (a to platí v menší míře dodnes) mnoho závěrů během několika let poopravil nebo dokonce i zcela pozměnil geometricky narůstající počet studovaných taxonů i souborů znaků. Při srovnání molekulární a „morfologické“ klasifikace vidíme význam molekulárních znaků zejména ve třech bodech:
• podporují předpokládanou monofyletičnost skupiny (na úrovni čeledí, jež nás bude zajímat nejvíce, je to kupodivu ve většině případů);
• dovolují rozhodnout mezi více hypotézami morfologické systematiky o zařazení skupin (např. u řádů Ericales či Urticales, v jejichž řazení se hlavní autoři morfologických klasifikací rozcházeli);
• dovolují zařadit skupiny, které pro svou biologickou či morfologickou specifitu byly velkým oříškem a v morfologických systémech se objevovaly často téměř chaoticky na různých místech, podle toho, jaký způsob argumentace autor zvolil (např. mnohé skupiny vodní, parazitické nebo masožravé).
Na druhou stranu je třeba zdůraznit, že objektivní molekulární „stromy“ zůstávají (a možná ještě dlouho zůstanou) nekompletní: Stále není dost podkladů pro přesné ukotvení některých taxonů (i některých nám dost známých čeledí, např. Boraginaceae), které tak zůstávají s nálepkou „unplaced“ nezařazeny.
Molekulární systematika a klasifikační hierarchie
Kladistika a její metodologii používající molekulární systematika se od počátku střetly se zavedenou hierarchickou strukturou taxonů na vyšších úrovních (řádových či nadřádových); tam bylo zcela běžné užívání parafyletických taxonů, dovolujících pojímat a seřazovat taxony do několika málo specifikovaných kategorií (oddělení, třída, podtřída, řád popř. nadřád). Princip striktní monofylie by navyšoval počet hierarchických taxonomických kategorií do závratných počtů, a tak se zejména anglosaští badatelé vydali cestou neformální klasifikace velkých skupin. Při tomto způsobu je např. u krytosemenných rostlin nejvyšší formalizovanou kategorií řád (Fabales apod.) a základní štěpné větve jsou označovány různě, nejčastěji termínem vycházejícím z podtříd morfologické klasifikace (Rosidae, Asteridae), obvykle však nelatinizovaným (Rosids – rosidy, Asterids – asteridy) a u velkých skupin dále štěpeným na číslované skupiny (např. Eu-rosids I, Eu-rosids II apod.). Tento způsob dovoluje názvy skupin počešťovat (rosidy). Praktická botanika pracuje zejména s kategoriemi čeledí, nejvýše řádů, a proto se s těmito skupinami obvykle nebude setkávat; v souborných dílech typu určovacích klíčů nebo učebnic se však začínají objevovat.
Objektivizace „systému“ v literatuře, na webu i v učebnicích
Nesmírný rozmach získávání molekulárních dat a molekulární systematiky jako takové vedl na konci minulého století k vytvoření skupiny vědců „Angiosperm Phylogeny Group“, jež se pokusila dát dohromady objektivní výčet řádů a čeledí krytosemenných rostlin a publikovala jej (APG I 1998). Skupina si zvolila za „domovskou“ adresu americkou Missouri Botanical Garden, a tak není divu, že na stránkách této zahrady se počátkem našeho století objevil od té chvíle nejdůležitější internetový zdroj zabývající se klasifikací krytosemenných rostlin – Angiosperm Phylogeny Website (Stevens et al. 2001), doplňovaný průběžně a představující asi nejčerstvější pohled na stav bádání v této oblasti. Skupina APG pak publikovala v r. 2003 (APG II 2003) krytosemenný seznam inovovaný podle výsledků pětiletého bádání a publikaci podobného typu lze v dalších létech opakovaně očekávat. Fylogenetické klasifikaci všech skupin vyšších rostlin (včetně mechorostů) a současně vyhynulých skupin (!) bylo v nedávné době věnováno souborné číslo renomovaného časopisu American Journal of Botany (2004); tento soubor kritických review, dostupný i na webu, bych vřele doporučil vážným zájemcům o problematiku. Z hlediska taxonomických kategorií je nutno zdůraznit, že uvedený kodifikující systém užívá v mnohých případech mnohem širší pojetí řádů a čeledí, než na jaké jsme ve středoevropské systematice zvyklí (konkrétní příklady budou uvedeny u pravých dvouděložných).
Učebnice a příručky zařazují nové poznatky obvykle ve druhém sledu. První učebnice zohledňující jednoznačně molekulární data byla publikována v Americe v r. 1999 (Judd et al. 1999) a záhy se dočkala druhého vydání (Judd et al. 2002). Mezi oběma vydáními je vidět veliký pokrok v hledání „uchopitelných“ znaků, podporujících a charakterizujících molekulárními znaky vylišené taxony. Uživatelsky příjemnou učebnicí je však až bohatě ilustrovaná učebnice Simpsonova (Simpson 2006). Je potěšitelné, že ve stejném roce vycházejí na totéž téma na Slovensku stručná, leč výstižná vysokoškolská skripta ve slovanském jazyce (Mártonfi 2006).
Mnohé nově pojímané skupiny v molekulární klasifikaci však stále na své charakteristické nápadné znaky čekají. Proto není divu, že se jim brání určovací příručky; i zde se však objevuje první vlaštovka v 93.(!) vydání Schmeil-Fitschenovy Flóry Německa (Seybold 2006).
Změny u kapraďorostů
Pomineme-li některé obtížně zařaditelné vymřelé skupiny (jimiž se zde nezabýváme), byly dlouhodobě rozeznávány 3 resp. 4 různě velké větve – známé plavuně, přesličky a kapradiny, vesměs s bohatým a často velmi dlouhým vývojem v minulosti, a drobná skupina tropických rodů Psilotum, Tmesipteris (Psilotopsida), známá prakticky pouze z recentu. Zcela samostatný vývoj plavuní (Lycopodiophyta), oddělivších se od předků všech dalších větví cévnatých rostlin, ukázala již dávno paleobotanika a podpořily ji i kladistické studie (Kenrick et Crane 1997).
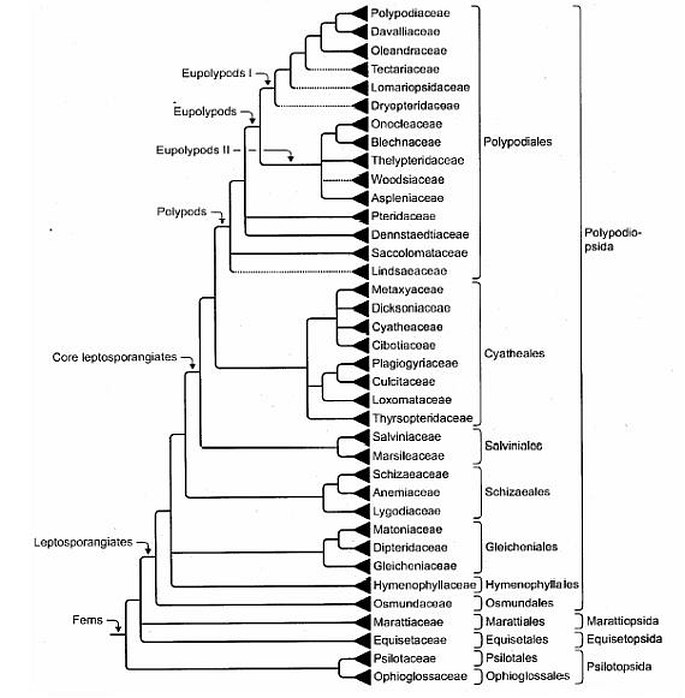
Molekulární systematika kapraďorostů „startuje“ v r. 1995 (Pryer et al. 1995) a tehdy zjištěné vztahy v průběhu dalších let prakticky jen zpřesňuje (nejnovější přehled viz Smith et al. 2006). Hlavní zjištění můžeme shrnout do následujících bodů (obr. 1):
• Přesličky a kapradiny jsou si příbuznější, než se předpokládalo a tvoří spolu s morfologickou třídou Psilotopsida jednotnou vývojovou větev nazývanou Moniliophyta (moniliofyty);
• Rozpadla se skupina eusporangiátních kapradin: drobné Ophioglossales vyskytující se i u nás (např. Botrychium) a mohutné tropické Marratiales nejsou blízce příbuzné;
• Ophioglossales tvoří jednu vývojovou větev s Psilotales (zejména na základě znaků v gametofytní generaci), skupinou považovanou doposud za zcela samostatnou;
• Moniliophyta (= kapraďorosty bez plavuní) jsou tedy reprezentovány 4 monofyletickými skupinami, jimž je ponechán taxonomický rank tříd: Psilotopsida, Equisetopsida, Marattiopsida a Polypodiopsida (= leptosporangiátní kapradiny).
Vlastní systém leptosporangiátních kapradin nedoznal zásadnějších změn. Velká čeleďová diverzifikace řádu Polypodiales (do níž náleží většina našich kapradin), užitá např. již v Květeně ČR (Hejný et Slavík 1988), byla molekulární systematikou potvrzena. Sluší se podotknout, že „otcem“ této klasifikace, založené zejména na anatomii řapíku a vřetene čepele, je Karel Bořivoj Presl, jakkoliv jeho práce byly na více než sto let oslyšeny. Zajímavé je ještě zjištění, že tzv. vodokapradiny (heterosporická skupina čeledí Salviniaceae, Marsileaceae) tvoří sice samostatnou vývojovou větev, zdaleka však ne tak izolovanou ostatním kapradinám, jak předpokládala morfologická systematika.
Obr. 1 – Kladogram skupiny Moniliophyta (Smith et al. 2006)
Problematika nahosemenných
Vymezení hlavních vývojových linií recentních nahosemenných se zdá jednoznačné (mám na mysli skupiny cykasů – Cycadophyta, jinanů – Ginkgoophyta, jehličnanů – Pinophyta a liánovců – Gnetophyta). Revoluční úvahy paleobotaniků o dvou vývojových liniích nahosemenných, „manoxylické“ (z recentních Cycadophyta) a pyknoxylické (ostatní skupiny), založené na anatomii dřeva, souměrnosti semen a dalších znacích (Beck 1988, Stewart et Rothwell 1992), nedožily konce minulého století. Monofylie nahosemenných se tedy jeví jednoznačně, překvapivá je však nesamostatnost liánovců (obr. 2), které v mnoha molekulárních studiích vycházejí jako součást větve jehličnanů s nejbližší příbuzností k našim borovicovitým (Pinaceae) (odkazy viz Simpson 2006).
Méně překvapivá je vzájemná provázanost i morfologicky blízce příbuzných čeledí Cupressaceae a Taxodiaceae (řazených i v minulosti do jednoho řádu), které se v klasické rodové náplni jeví jako parafyletické taxony, a proto jsou v novém pojetí součástí široké čeledi Cupressaceae; zcela izolovanou větev ale představuje japonský keř Sciadopitys verticillata, který si tak „zasloužil“ vlastní čeleď Sciadopityaceae.
Nejasnosti ve vztazích nahosemenných přetrvávají na úrovni vymřelých skupin, ty se však vymykají z rámce našich úvah.
Obr. 2 – Základní kladogram nahosemenných (Simpson 2006)

Krytosemenné (Magnoliophyta, Angiospermae) a jejich základní schéma
Fylogenetický strom krytosemenných byl vždy nejsložitější a vzhledem k jejich relativnímu geologickému mládí byl také nejméně podporován výsledky paleobotanických bádání. Po většinu dvacátého století uznávala morfologická evoluční klasifikace základní diversifikaci na dvouděložné a jednoděložné a za nejprimitivnější krytosemenné rostliny pak v rámci dvouděložných okruh řádu Magnoliales (ve shodě s převládající euanthiovou teorií); u tříd jednoděložných a dvouděložných pak předpokládala diverzifikaci na několik velkých podtříd – v ní se poněkud lišily obě základní koncepce (Cronquist 1981, Tachtadžjan 1977 – viz obr. 3). Velmi obecně možno říci, že molekulární systematika v průběhu posledních 14 let:
• rozbořila klasické členění na jedno- a dvouděložné;
• dospěla k závěru, co je nejpůvodnější krytosemennou rostlinou;
• potvrdila diverzifikaci odvozených skupin s určitou podobností odpovídající (i označením) podtřídám morfologického systému;
• naopak na bázi každé skupiny existuje obvykle několik samostatných větví obsahujících po jedné (obvykle) čeledi, které nejeví bližší příbuznost ke zbytku skupiny;
• ukázala, že nejsložitější je míra příbuznosti a zobrazení vzájemných vztahů u zcela bazálních větví krytosemenných.
Vývoj základní diverzifikace ukazuje Tab. 2. Zatímco jednoděložné a pravé dvouděložné (ve zřídka používaném latinském označení Rosopsida) jsou monofyletickými skupinami, byť v případě dvouděložných je tato monofylie podporována jediným „morfologickým“ znakem – trikolpátním pylem, skupina bazálních dvouděložných (či možná lépe bazálních krytosemenných) soustřeďuje větší počet často máločeleďových větví a její označení je pragmatické a nepoužitelné ze striktně fylogenetického hlediska. Kdybychom sledovali vývoj kladogramů v raných létech molekulární systematiky, viděli bychom, že na postu bazální čeledi se vystřídalo několik „kandidátů“ (např. Ceratophyllaceae, Chloranthaceae), jejichž zařazení je i dnes sporné, než se do kotle molekulárních analýz dostala monotypická novokaledonská čeleď Amborellaceae; Keř Amborella trichopoda pak vstoupil do 21. století jako nejprimitivnejší krytosemenná rostlina, následován dvěma řády Nymphaeales a Austrobaileyales (kam ze známějších čeledí patří Illiciaceae), u nichž byly zjištěny i další velmi primitivní 1) znaky (viz tab. 2). Tato skupina získala ve fylogenetické literatuře i neformální označení ANITA (group) (viz např. Mártonfi 2006). Následné jádro bazálních krytosemenných pak tvoří magnolidy, skupina řádů v podstatě odpovídající Tachtadžjanově morfologické nejprimitivnejší podtřídě dvouděložných Magnolidae (Magnoliales, Laurales, Canellales, Piperales).
Z hlediska praktického uživatele systému je tento výsledek studia v podstatě kladný; následující velké skupiny jednoděložných a pravých dvouděložných získávají mnoho nápadných (téměř) výlučných znaků, které jsou vlastně společné pro bazální krytosemenné a jednoděložné oproti pravým dvouděložným (např. pseudomonokotylie, častá trojčetnost květů apod.).
__________________________________________________
Tab. 2 – Základní členění krytosemenných
• Až do 1993 (morfologická systematika):
Dvouděložné (Magnoliopsida) – jednoděložné (Liliopsida)
• 1993-ca 2000 (raná molekulární systematika):
Bazální dvouděložné (Magnoliopsida) – jednoděložné (Liliopsida) – pravé dvouděložné (Rosopsida)
• 21. století (molekulární systematika opouští plenky…):
Pre-angiospermy (skupina ANITA) + magnoliová větev (magnolidy) = dohromady „bazální krytosemenné“ – jednoděložné (monocots) – pravé dvouděložné (Eu-Dicots)
• ANITA – parafyletická skupina s mnoha starobylými znaky, nejdůležitější: nedokonalý konduplikátní plodolist: není okraji srostlý, pouze slepený; 4buněčný zárodečný vak!
__________________________________________________
Obr. 3 – Schematické znázornění a stručná charakteristika podtříd podle Tachtadžjana (černý podklad) a širší pojetí Cronquista (v šedých elipsách)

Jednoděložné (monocots, Liliopsida) včera a dnes
Změny v nazírání na velikost a pojetí čeledí u jednoděložných probíhaly kontinuálně od osmdesátých let (zejména u tzv. liliid) a molekulární systematika jim, jakkoliv někdy v jiném postavení na fylogenetickém stromu, dala ve většině případů za pravdu. To platí i o základní diverzifikaci na dřívější podtřídy Alismatidae, Liliidae, Commelinidae a Arecidae, kde molekulární systematika jen zfúzovala poslední dvě skupiny v commelinidy s palmami na bázi. Rozčlenění entomogamní skupiny liliidů na mnoho čeledí v rámci dvou řádů Asparagales a Liliales (nápadnými vnějšími morfologickými znaky prakticky nerozlišitelných) je užíváno od vydání přehledové příručky o jednoděložných (Dahlgren, Clifford, Yeo 1985 – viz též tab. 3 a 4), a proto se s ním setkáváme i v našem novém určovacím Klíči (Kubát et al. 2002); Objevitelem většiny rozlišovacích znaků a autorem základního schematu byl však již o 16 let dříve Huber (1969) v obsáhlé, leč německy psané práci o anatomii semen. Jediným zásadním přesunem molekulární systematiky z hlediska středoevropské flóry byl rozpad čeledi Melanthiaceae, z níž Tofieldia a příbuzné rody „odputovaly“ až do následně diskutované skupiny primitivních alsimatidů a vytvořily zde samostatnou čeleď Tofieldaceae.
Nejnápadnější změny posledního desetiletí tak můžeme zaznamenat opět u nejpůvodnější skupiny alismatidů:
• bazální (opět monotypickou) čeledí jednoděložných jsou Acoraceae; puškvorec soustřeďuje některé znaky, které se nevyskytují u jednoděložných vůbec nebo zcela výjimečně (tvorba silic, perisperm apod.), a svou bývalou čeleď Araceae připomíná jen palicovitým květenstvím, ale už ne dokonalou oboupohlavnou stavbou trojčetných květů a absencí toulcovitého listenu;
• naopak následující Araceae „pohltily“ okřehkovité (Lemnaceae), což je při pohledu na redukované květy okřehků (které se v mirném pásu netvoří) a přechodný vzplývavý druh Pistia stratiotes při troše fantazie pochopitelné;
• drobné, převážně vodní a bažinné čeledi vlastních alismatidů nedoznaly větších změn; překvapivé je snad jen spojení nenápadně kvetoucích řečanek (Najadaceae) s rody čeledi Hydrocharitaceae.
Tab. 3 – Hlavní rozlišovací znaky mezi řády Asparagales a Liliales

Tab. 4 – Zastoupení našich domácích (tučně) a běžně pěstovaných rodů v čeledích nového systému „liliovitých“ jednoděložných

Hlavní skupiny pravých dvouděložných (eu-dicots, Tricolpatae, Rosopsida)
Bazální diverzifikace této skupiny obsahuje opět množství drobných skupin, promíšené většími skupinami odpovídajícími v podstatě podtřídám morfologické (Tachtadžjanovské) klasifikace. Na ně pak navazuje zásadní větvení většiny dvouděložných na skupiny Rosidae (rosidy) a Asteridae (asteridy) (obr. 5). Bodově lze komentovat bazální diverzifikace následovně:
• bazální skupina Ranunculidae je zrcadlovým obrazem morfologické podtřídy téhož jména, vylišené Tachtadžjanem (kterému se tak oproti Cronquistovi podařilo intuitivně vystihnout výrazný rozdíl a nikoli návaznost mezi převážně dřevinnými magnolidy a převážně bylinnými ranunculidy);
• následuje „hřebenový kladogram“ několika převážně tropických nebo jihopolokoulových samostatných čeledí nebo drobných skupin, morfologicky velmi málo podobných. Klasickým příkladem je skupina tzv. proteidů (často uváděná jako řád Proteales), spojující do jedné skupiny Proteaceae (s jihopolokoulovým tzv. gondwanským areálem), reliktní temperátní Platanaceae a vodní převážně tropické Nelumbonaceae;
• druhou velkou a kompaktní skupinou jsou caryophyllidy (Caryophyllidae), ve složení uznávaném z větší části již morfologickou systematikou; molekulární poznatky sem však přivanuly ještě skupinu masožravých čeledí řádu Droserales.
Obr. 5 – Fylogenetické vztahy v rámci pravých dvouděložných (Judd et Olmstead 2004). Použito nových termínů pro velké skupiny v rámci rosidů a asteridů

Tato karnivorní skupina je klasickým příkladem prokázané příbuznosti biologicky podobných, avšak květní morfologií a dalšími znaky odlišných skupin, které byly ve všech morfologických systémech řazeny zcela vzdáleně (Nepenthaceae vs. Droseraceae). A poslouží nám ještě jako jeden příklad: zcela proti převládajícímu trendu velkých (protože nutně monofyletických) čeledí zde molekulární systematika potvrzuje jako oprávněnou jednu z mála čeledí popsaných českými autory – Drosophyllaceae (Chrtek, Slavíková et Studnička 1989).
Kladogram rosidů a asteridů kopíruje svou strukturou základní kladogram pravých dvouděložných: na bázi několik menších skupin, následovaných hlavní „vidličkou“ na skupiny: rosidy I (fabidy) a II (malvidy), asteridy I (lamiidy) a II (asteridy) (cf. obr. 5). Zatímco pro rozlišení obou skupin asteridů existuje značná podpora morfologických znaků (ne nadarmo jejich jádro tvoří tachtadžjanovské podtřídy Lamiidae a Asteridae), jako např. převládajíci typ postavení semeníku, tvorba odlišných sekundárních metabolitů nebo v poslední době typ sympetalie (viz dále), morfologické odlišení obou hlavních skupin rosidů zatím uspokojivě nalezeno nebylo.
Čeleďová problematika a příklady změn
Je pochopitelné, že u pravých dvouděložných došlo při studiu pomocí molekulární systematiky k největšímu množství změn a jejich podrobné komentování by zdaleka přesáhlo rámec tohoto sborníku. Proto jsou dále uvedeny jen 3 příklady změn v pojetí příbuzných čeledí, dotýkající se bezprostředně naší plané nebo u nás běžně pěstované květeny.
(a) Malvales – Malvaceae
Čtenář středoevropských příruček je zvyklý na řád složený z čeledí Tiliaceae, Sterculiaceae (Theobroma), Bombacaceae (Gossypium, Adansonia) a Malvaceae. Toto tradiční členění však poznatky molekulární systematiky zcela rozložily (obr. 6). Jak ukazuje tab. 5, ani jedna z dosavadních čeledí nezůstala ve své původní podobě; jako zcela heterogenní se ukázaly zejména první dvě výše uvedené čeledi, kde se ukázalo, že tyto čeledi obsahovaly jak velmi primitivní skupiny (v tab. 5 podčeledi Byttnerioideae a Grewioideae), tak větší počet skupin pokročilejších. V tabulce jsem se pokusil vystihnout i určité korelace tohoto pojetí s vývojem některých nápadných morfologických znaků od primitivních po pokročilé (volnost → srostlost tyčinek, klesající počet funkčních prašných váčků, typ gynecea). Jak je však patrno, struktura znaků jeví i v této zjednodušené podobě značně retikulátní strukturu a dává tak za pravdu novému pojetí; s lítostí je tak nutno konstatovat, že v nejbližších určovacích klíčích bude národní strom Tilia náležet do čeledi slézovitých. A pokud se setkáme s řádem Malvales, bude obsahovat mnohem více čeledí, byť i morfologickou systematikou uznávaných jako slézovitým vzdáleněji příbuzné (Dipterocarpaceae, Cistaceae, Thymelaeaceae apod.)
Obr. 6 – Fylogenetické vztahy v rámci široce pojímané čeledi Malvaceae (Judd et. al. 2002)

(b) Ericales – Ericaceae
Ericales jsou podobný případ: řád v původním pojetí vplul do ranku čeledi a skupiny se promíchaly (obr. 6, tab. 6). Vzácně zůstaly v původní šíři (Vacciniaceae resp. Vaccinioideae), svým způsobem i Pyrolaceae a saprofytické Monotropaceae, naopak se rozpadla původní úzce pojatá čeleď Ericaceae; hlavním morfologickým znakem podporujícím molekulární data se zde stal ontogenetický znak tzv. pozdní resp. časné inverze prašníků, rozdělující fylogeneticky původnější a pokročilejší skupiny. Pozorného čtenáře jistě zaujme, že do výrazně zúženého příbuzenství vřesovců a rododendronů přibyly rody původní čeledi Empetraceae, oddělované od zbytku vřesovcovitých na základě zcela jiných počtů květních částí (obecně méně četné a počty ne zcela pravidelné květy); jednou z možností těchto květních odlišností je anemogamie šich oproti zbytku vřesovcové větve, vlastnost, která má obecně v evoluci za následek redukci v květech.
Obr. 6 – Fylogenetické vztahy v rámci široké čeledi Ericaceae

(c) Scrophulariaceae a příbuzné čeledi
Příklad nejzapeklitější a pro temperátní čeledi ojedinělý: totální rozpad „zavedené“ čeledi, signalizovaný již v roce 1995 (Olmstead et Reeves 1995), obecně potvrzovaný a akceptovaný (a spíše čím dál více dokonávaný – Olmstead et al. 2001). Na rozdíl od předcházejících příkladů jsou nově vzniklé čeledi obsahující části původních krtičníkovitých
• od sebe značně fylogeneticky vzdáleny a „promíšeny“ jinými příbuznými čeleděmi monofyletického řádu Lamiales (obr. 7);
• a naopak v případě čeledi Plantaginaceae v novém pojetí zahrnují zcela morfologicky i biologicky nepodobné skupiny rostlin.
Pomineme-li zatím ne zcela uznávané čeledi Phrymaceae (Mimulus) či Linderniaceae (Lindernia, Limosella) a zastavíme se u třech největších čeledí Scrophulariaceae s. str., Orobanchaceae a Plantaginaceae, musíme konstatovat, že určitá i nemolekulární podpora tu existuje. Rody zbylé v krtičníkovitých (z našich Verbascum, Scrophularia) se vyznačují zcela specifickým typem prašníků se srostlými prašnými váčky, který se u ostatních dvou čeledí nevyskytuje. Orobanchaceae v novém pojetí soustřeďují „pod jednu střechu“ všechny parazity a poloparazity, vyznačující se kromě této biologické vlastnosti i speciální fytochemií (alkaloid orobanchin). Černý Petr téměř nemožné charakterizovatelnosti tak zůstává výše zmíněné čeledi Plantaginaceae, v níž najdeme kromě zbytku autotrofních rodů krtičníkovitých bývalé Plantaginaceae s.str., Globulariaceae a navíc vodni Callitrichaceae a Hippuridaceae. Tady je klasické pátrání po morfologické příbuznosti prací více než sisyfovskou.
Obr. 7 – Fylogenetické vztahy širokého řádu Lamiales (Judd et al. 2002); větve řazené tradiční systematikou do čeledi Scrophulariaceae jsou označeny hvězdičkou

Tab. 5 – Nové pojetí čeledi Malvaceae s. latissimo (incl. Tiliaceae, Sterculiaceae, Bombacaceae): její členění a charakteristika jednotlivých podčeledí


Tab. 6 – Ericaceae s.l. – současné členění


Kladogramy a nové „morfologické“ znaky
Jak už jsem zmínil, první kladogramy, které vznikly při molekulárních analýzách, obvykle postrádaly podporu uchopitelných morfologických znaků. Příklady kladogramů z Juddovy učebnice i dalších prací v tomto příspěvku ukazují, že v tomto směru byl učiněn značný pokrok. Ten je často podporován dalším přístrojovým či metodologickým rozmachem podpůrných disciplín, studujících struktury a funkci rostlinného těla. Jedním z takových objevů je již zmíněné zjištění časného resp. pozdního obracení prašníků při ontogenezi květů vřesovcovitých rostlin (Kron et al. 2002), které je výsledkem časosběrného studia květních orgánů pomocí SEM.
Ještě nápadnějším příkladem zjištěným pomocí SEM, je tzv. časná a pozdní sympetalie u asteridů. Srostlá koruna je známým typickým znakem obou hlavních skupin, Asteridů I (lamiidů – řádů Gentianales, Solanales, Lamiales) a asteridů II (campanulidů – řádů Apiales, Dipsacales, Asterales); ukázalo se však, že u první skupiny se jednotlivé korunní lístky zakládají jako samostatná primordia a teprve poté srůstají dohromady (pozdní sympetalie), zatímco u asteridů II se již zakládají jako kruh s později se prodlužujícími 5 korunními cípy (časná sympetalie) (Leins 2000, Leins et Erbar 2003). Možná proto nám odkvétající koruna hluchavky zůstane snadno v ruce, zatímco koruna zvonku nikoli. Tento znak je pravděpodobně nejlepším nemolekulárním znakem odlišujícím tyto dvě větve.
Regionální pohled – co se změnilo ve skupinách středoevropské flóry?
Nástup molekulární systematiky, probuzení spící panny „zavedené“ taxonomické klasifikace, geometrická řada příspěvků zpracovávajících obrovské množství sekvenovaného materiálu (ono pověstné „Jáchyme, hoď ho do stroje“) následovaná syntetizujícími pracemi – z toho se vejde na několik stránek příspěvku jen několik zobecnění a několik výše uvedených příkladů. Jako přídavek si dovoluji uvést stručný přehled konkrétních změn ve střední Evropě (tab. 7). Z tabulky vyplývá, že taxonomický dopad výsledků molekulární systematiky na běžně užívaných úrovních čeledí resp. rodů se může projevit v podstatě trojím způsobem:
• fúzí čeledí
• „rozdrobením“ čeledi
• přesunem rodu (nebo několika rodů) do jiné čeledi
Souhrn
Výsledky srovnání morfologické a molekulární klasifikace (dovolíme-li si zjednodušující nadhled) lze shrnout do následujících bodů:
• V hrubém zrnu se obě klasifikace zásadně neliší: „dálkový“ přenos čeledí (alespoň temperátních) proběhl většinou u takových, jejichž zařazení bylo často složité zdůvodňovat (paraziti, vodní, masožravé, malé čeledi dřevin)
• Jemné zrno často ukazuje na zcela jiné vztahy v rámci jinak dobře ohraničené skupiny (morfologického řádu) – viz Malvaceae, Ericaceae a další příklady
• Totální „reorganizace“ čeledi či řádu je však vzácná (Scrophulariaceae)
• Tendence je spíše k vytváření velkých čeledí (na úrovni morfologických řádů), ale i naopak (nová čeleď Drosophyllaceae)
• Kladogramy jsou „naplňovány“ znaky, často novými a nezřídka ontogenetickými (časná x pozdní inverze prašníků, časná x pozdní sympetalie)
Tab. 7 – Nejdůležitější změny pojetí čeledí či zařazení rodů ve středoevropské flóře


Na závěr: trochu oleje do ohně aneb jak nasadit brouka do hlavy
Jsme ve chvíli, kdy molekulární systematika a z ní vycházející fylogenetická klasifikace cévnatých opustily stránky impaktových časopisů a vtrhly do učebnic, příruček a určovacích klíčů; přání, jež vyslovili v souhrnném čísle časopisu American Journal of Botany věnovanému fylogenezi Judd et Olmstead (2004), se naplnilo neobvykle brzy. Věda je ale šťouravá, a tak není divu, že proti jednotné frontě monofyletické kladistiky jako jediné „pravé“ klasifikační metody se zdvihá mírná polemika. O srovnání kladistické a evoluční (fyletické) klasifikace (a o právo druhé metodologie na místo na slunci) se pokoušejí mnozí – doyen evolucionistů stoletý zoolog Ernst Mayr (Mayer et Bock 2002) i příslušníci střední generace; zodpovědně polemizuje např. Hörandl (2006) v časopise Taxon. I když nepochybuji o obrovském přínosu molekulární systematiky pro taxonomii jako takovou, z hlediska „utilitární“ botaniky mne občas napadne: nepřehodili jsme výhybku na cestu k moderním výšinám příliš brzy?
___________________________________________________
Poznámky
1) Primitivní a pokročilé = odvozené znaky je dnes zvykem označovat mezinárodními kladistickými termíny plesiomorfni a apomorfní; vzhledem k předpokládáné čtenářské obci jsem se rozhodl používat původní, pochopitelnější označení.
___________________________________________________
Literatura
American Journal of Botany (2004): Vol. 91(10) /Speciální číslo věnované fylogenezi cévnatých rostlin/
APG I /Angiosperm Phylogeny Group/ (1998): An ordinal classification for the families of flowering plants.- Ann. Missouri Bot. Gard. 85: 531-553.
APG II /Angiosperm Phylogeny Group/ (2003): An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. – Bot. J. Linnean Soc. 141: 399-436.
Beck G. (1988): Origin and evolution of Gymnosperms. – Columbia Univ. Press., New York
Chase, M. W. et al. (1993): Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. – Ann. Missouri Bot. Gard. 80: 528-580.
Chrtek J., Slavíkova Z., Studnička M. (1989): Beitrag zur Leitbündelanordnung in der Kronblättern von ausgewählten Arten der fleischfressenden Pflanzen. – Preslia 61: 107-124.
Cronquist, A. (1981): An Integrated System of Classification of Flowering Plants. Columbia Univ. Press, New York.
Cronquist A. (1988): The evolution and systematics of flowering plants. Ed.2. – New York Bot. Garden.
Dahlgren R., Clifford, H. T., & Yeo, P. F. (1985): The Families of the Monocotyledons. – Springer Verlag, Berlin.
Hejný S. et Slavík B. /ed./ (1988): Květena ČSR. Vol. 1. – Academia Praha.
Hennig W. (1966): Phylogenetics Systematics. – Univ. Illinois Press, Urbana.
Hendrych R. (1977, 1986): Systém a evoluce vyšších rostlin. Ed. 1, 2. – SPN Praha.
Hörandl E. (2006): Paraphyletic versus monophyletic taxa – evolutionary versus cladistic classifications. – Taxon 55(3): 564-570.
Huber H. (1969): Die Samenmerkmale und Verwandschaftsverhältnisse der Liliiflorae. – Mitt. Bot. Staatsamml. München 8:219-538.
Judd W.S. et al. (1999): Plant Systematics. A phylogenetic approach. – Sinauer Inc., Sunderland, Mass.
Judd. W.S. et al. (2002): Plant Systematics. A phylogenetic approach. Ed. 2. – Sinauer Inc., Sunderland, Mass.
Judd. W.J., Olmstead, R. G. (2004): A survey of tricolpate (eudicot) phylogenetic relationships. – American J. Bot. 91: 1627-1644.
Kenrick P., Crane P. R. (1997): The origin and early diversification of land plants. – Smithsonian Institution, Washington.
Kron A. et al. (2002): A phylogenetic classification of Ericaceae: Molecular and morphological evidence. – Bot. Rev. 68: 335-423.
Kubát et al. (2002): Klíč ke květeně České republiky. – Academia Praha.
Leins P. (2000): Blüte und Frucht. Aspekte der Morphologie, Entwicklungsgeschichte, Phylogenie, Funktion und Ökologie. – E.Schweizerbart, Stuttgart.
Leins P. et Erbar C. (2003): Floral developmental features and molecular data in plant systematics. – In:Stuessy T. et al. /ed./, Deep Morphology – Toward a rennaisance of morphology in plant systematics, p. 81- 105. Gantner Verlag, Liechtenstein
Mártonfi P. (2006): Systematika cievnatých rastlín. Ed. 2 – Univ. P.J. Šafárika Košice /skripta/
Mayr E., Bock W.R. (2002): Classifications and other ordering systems. – J. Zool. Syst. Evol. Research 40:169- 194.
Nandi O.I., Chase, M. W., Endress, P. K. (1998): A combined cladistic analysis of angiosperms using rbcL and non-molecular data sets. – Ann. Missouri Bot. Gard. 85: 137-212.
Novák F.A. (1961, 1972): Vyšší rostliny. Ed. 1, 2. – Academia Praha.
Olmstead R.G., Reeves P.A. (1995): Evidence for the polyphylly of the Scrophulariaceae based on chloroplast rbcL and ndhF sequences. – Ann. Missouri Bot. Gard. 82:176-193.
Olmstead R.G. et al. (2004): Disintegration in the Scrophulariaceae. – Amer. J. Bot. 88(2): 348-361.
Pryer K. M., Smith A. R., & Skog J. E. (1995): Phylogenetic relationships of extant ferns based on evidence from morphology and rbcL sequences. – Amer. Fern J. 85: 205-282.
Seybold S. (2006): Schmeil-Fitschen´s Flora von Deutschland. Ed. 93. – Quelle et Mayer, Stuttgart.
Simpson M. (2006): Plants Systematics. – Elsevier, Amsterdam
Smith A.R. et al. (2006): A classification for extant ferns. – Taxon 55(3):705-731.
Soltis, D. E. et al. (2005): Phylogeny and Evolution of Angiosperms. Sinauer, Sunderland, Mass.
Stevens P. et al. (2001-): Angiosperm Phylogeny Website. –
http:// www.mobot.org/MOBOT/research/APweb
Stewart W., Rothwell G.W. (1992): Paleobotany and the evolution of the plants. –Cambridge Univ. Press.
Tachtadžjan A.A. (1987): Sistema magnoliofitovych. – Nauka Leningrad.